
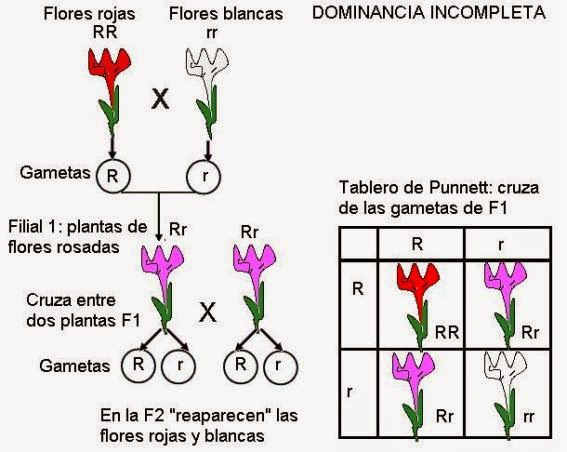
Dominancia incompleta
La
dominancia incompleta es una condición en la cual ningún alelo es dominante
sobre el otro. La condición es reconocida para heterocigotos que expresan un
fenotipo intermedio en relación a los fenotipos paternos. Si una planta roja se
cruza con una planta de flores blancas, la progenie será toda rosa. Cuando una
rosa se cruza con otra rosa, la descendencia es 1 roja, 2 rosas, y una blanca.

Alelos Múltiples.
Hablamos
de alelos múltiples cuando hay más de dos alelos alternativos posibles para
especificar ciertos rasgos.Un ejemplo típico lo constituyen los alelos del
sistema de grupos sanguíneos. Los alelos múltiples se originan de distintas
mutaciones en un mismo gen.Los 4 grupos sanguíneos: A, B, AB y O son resultado
de tres diferentes alelos de un sólo gen (iA, iB e iO), iA e iB son codominantes
sobre iO que es recesivo

En
algunos casos, el gen dominante no logra encubrir por completo la expresión del
gen recesivo, sino que ambos se expresan parcialmente. En los individuos
heterocigotas aparece entonces un tercer fenotipo, diferente del dominante y el
recesivo, e intermedio entre los dos. A estos tipos de herencia se les da el
nombre de “dominancia incompleta”. Por ejemplo, en ciertas plantas, las
flores pueden ser blancas, rojas o rosadas. El fenotipo “flores rosadas” se
observa en los individuos heterocigotas, como resultado de la expresión parcial
de los alelos que codifican los colores rojo y blanco. Cabe aclarar que, sin
embargo, la aparente “mezcla” solo se produce a nivel fenotípico, ya que los alelos
mantienen su individualidad, tal como lo advirtió Mendel. Así, de la cruza de
dos individuos rosados, se obtendrá una progenie 25% roja, 25% blanca y 50%
rosada.
En
el hombre, la hipercolesterolemia familiar es una enfermedad de origen
genético, caracterizada por altos niveles de LDL en sangre, lo que predispone a
la aterosclerosis y alto riesgo de enfermedad cardiovascular precoz. El
trastorno se debe a la herencia de un receptor de LDL anómalo, por lo que las
células no pueden endocitar estas lipoproteínas.


Ciertos
alelos son codominantes. Por ejemplo, en las personas existen tres
alelos (alelos múltiples) que codifican antígenos (también llamados
aglutinógenos) presentes en la membrana de los glóbulos rojos. El genotipo para
este rasgo determina el grupo sanguíneo (fenotipo) que presentará el individuo.
Como se puede observar en la siguiente tabla, los alelos IA e IB codifican los
antígenos A y B, respectivamente, y dominan sobre el alelo i, que codifica la
ausencia de antígeno. Los alelos IA e IB son codominantes. Esto resulta en la
expresión de ambos antígenos en la membrana de los glóbulos rojos, en los
individuos portadores de los dos alelos.
http://www.genomasur.com/BCH/BCH_libro/imagenescap_18/b.JPG
Se
denomina ligamiento a la herencia conjunta de los genes que están ubicados
en el mismo cromosoma. Dado que los cromosomas pasan completos a una de las
gametas durante la meiosis, los genes de un mismo cromosoma se heredan juntos.

Herencia
poligénica
Si
bien hasta ahora hemos analizado caracteres afectados por un solo par de genes,
muchas características fenotípicas son el resultado de la interacción entre
varios pares de genes.
Uno
de los patrones de herencia poligénica es el que se presenta en la herencia del
color de piel o de la estatura. Estos rasgos dependen de la influencia de
varios pares de genes, cuyo efecto es aditivo. Ambos son casos de herencia
cuantitativa, puesto que el fenotipo resulta de la cantidad de alelos
dominantes y recesivos heredados en todos los pares de genes que determinan el
rasgo.
Por
ejemplo, si un rasgo cuantitativo está determinado por tres pares de genes,
dicho rasgo alcanzará su máxima expresión fenotípica en el individuo con el
genotipo AACCBB, y la mínima, en el individuo con el genotipo aabbcc. A su vez,
el mismo fenotipo puede ser determinado por varios genotipos distintos (por
ejemplo: AaBBCc y AABbCc).En estos patrones de herencia, la población presenta
una variación fenotípica continua, cuya distribución de frecuencias da una
típica curva de Gauss.

Rasgos
autosómicos, ligados al sexo y holándricos
El
cariotipo humano está constituido por 44 cromosomas somáticos y 2 cromosomas
sexuales. El par sexual es el que determina el sexo cromosómico.Se denomina rasgo
autosómico a aquél que se halla determinado por genes ubicados en los
autosomas. La herencia de estos rasgos es independiente del sexo del individuo,
por cuanto los autosomas son los mismos en el varón y en la mujer.La herencia
de los genes ubicados en el par de cromosomas sexuales sí se ve afectada por el
sexo del individuo, ya que las mujeres heredan el par XX y los varones, el par
XY.
Los
cromosomas X e Y son homólogos solamente en pequeñas regiones, ubicadas en los
extremos, las cuales reciben el nombre de “regiones pseudoautosómicas”. Éstas
son las que permiten el apareamiento del X y el Y durante la meiosis que lleva
a la formación de los espermatozoides en los varones. Sin embargo, las
restantes porciones de los cromosomas sexuales no son homólogas, y por lo tanto
llevan genes distintos. A estas porciones se las denomina “heterólogas”.
Los rasgos determinados por los genes ubicados en las porciones heterólogas del
cromosoma X se conocen como “rasgos ligados al sexo” o “ligados al X”.
Las mujeres heredan una copia materna y otra paterna de dichos genes, pudiendo
resultar homocigotas dominantes, homocigotas recesivas o heterocigotas para
dichos rasgos. En cambio, los varones solo heredan una copia del gen,
proveniente de su madre, por lo que sus genotipos pueden ser hemicigota
dominante o hemicigota recesivo. Los genes ubicados en las porciones
heterólogas del cromosoma Y son los “genes holándricos” o “ligados al
Y”. Éstos son exclusivos de los varones.

Los
cromosomas sexuales
El
cromosoma X es submetacéntrico. La mayoría de los genes de los cromosomas
sexuales se encuentran en el cromosoma X (se calcula que porta entre 1.500 y
2.500 genes). De allí que la expresión “genes ligados al sexo” se haya
utilizado tradicionalmente para referirse a los genes ubicados en dicho
cromosoma.
En 1949, Murray Barr observó que en las células de las hembras, aparecía un corpúsculo nuclear intensamente teñido, al que se conoce desde entonces como corpúsculo de Barr. En 1961, Mary Lyon establece que el corpúsculo de Barr es uno de los cromosomas X, que se presenta como heterocromatina (heterocromatina sexual).
En las primeras semanas de desarrollo, en las células embrionarias, uno de los cromosomas X se inactiva. Cuál de ellos lo haga, es azaroso. Sin embargo, una vez inactivado un X, todas las células descendientes mantienen inactivo el mismo cromosoma. Así, el embrión resulta formado por clones de células que expresan el cromosoma X materno y otros que expresan el cromosoma X paterno. Por lo tanto, las hembras son mosaicos en lo que respecta a la expresión del cromosoma X.
En 1949, Murray Barr observó que en las células de las hembras, aparecía un corpúsculo nuclear intensamente teñido, al que se conoce desde entonces como corpúsculo de Barr. En 1961, Mary Lyon establece que el corpúsculo de Barr es uno de los cromosomas X, que se presenta como heterocromatina (heterocromatina sexual).
En las primeras semanas de desarrollo, en las células embrionarias, uno de los cromosomas X se inactiva. Cuál de ellos lo haga, es azaroso. Sin embargo, una vez inactivado un X, todas las células descendientes mantienen inactivo el mismo cromosoma. Así, el embrión resulta formado por clones de células que expresan el cromosoma X materno y otros que expresan el cromosoma X paterno. Por lo tanto, las hembras son mosaicos en lo que respecta a la expresión del cromosoma X.

El
hecho de que en el sexo XX, cada célula exprese uno solo de estos cromosomas,
permite lo que se denomina “compensación de la dosis”. Esto significa
que los productos codificados por los genes presentes en el X se expresan en la
misma dosis que en los machos, portadores de una sola versión del cromosoma. Es
decir que, funcionalmente, las hembras son hemicigóticas para los genes ligados
al X. Incluso en los individuos que tienen un número anormal de cromosomas
sexuales, como en el síndrome de Klinefelter (XXY) o Turner (X0) se cumple la
regla según la cual se inactivan todos los X menos 1.

La
herencia citoplasmática
Las
mujeres heredan los cromosomas X de su madre y de su padre. Los varones heredan
el cromosoma X de su madre y el Y de su padre. Pero todos heredamos el
cromosoma mitocondrial de nuestra madre. En las células animales, existe una
herencia citoplasmática además de la nuclear: el ADN mitocondrial. Esta
herencia se produce por vía exclusivamente materna, pues como ya vimos al
tratar el tema fecundación, es el óvulo el que aporta el citoplasma, con sus
organoides, para formar la cigota.
La comparación del ADN mitocondrial posibilita el estudio de un linaje estableciendo los parentescos por línea materna. Este método se ha empleado en la búsqueda de nuestros ancestros como especie y también para resolver algunos enigmas particulares. Por ejemplo, se logró confirmar la identidad de los restos de la última emperatriz rusa, Alejandra, al comparar su ADN mitocondrial con el de su sobrino nieto por vía materna, el príncipe Felipe de Edimburgo (historia narrada en el libro “Una tumba para los Romanov”, de Raúl Alzogaray).
La comparación del ADN mitocondrial posibilita el estudio de un linaje estableciendo los parentescos por línea materna. Este método se ha empleado en la búsqueda de nuestros ancestros como especie y también para resolver algunos enigmas particulares. Por ejemplo, se logró confirmar la identidad de los restos de la última emperatriz rusa, Alejandra, al comparar su ADN mitocondrial con el de su sobrino nieto por vía materna, el príncipe Felipe de Edimburgo (historia narrada en el libro “Una tumba para los Romanov”, de Raúl Alzogaray).
Tabla
1: Ejemplos de rasgos humanos monogénicos
|
||||||||
Característica
|
Carácter
dominante
|
Carácter
recesivo
|
||||||
Nacimiento
del cabello en la frente
|
Pico
de viuda
|
Recto
|
||||||
Lóbulo
de la oreja
|
Separado
|
Adherido
|
||||||
Pulgar
|
Recto
|
Curvo
|
||||||
Entrecruzamiento
de los pulgares
|
Izquierdo
sobre derecho
|
Derecho
sobre izquierdo
|
||||||
Hoyuelo
en la barbilla
|
Presencia
|
Ausencia
|
||||||
Pecas
|
Presencia
|
Ausencia
|
||||||
Color
del pelo
|
Oscuro
|
Claro
|
||||||
Textura
del pelo
|
Rizado
|
Lacio
|
||||||
Pigmentación
|
Normal
|
Albinismo
|
||||||
Hipertricosis
de la oreja (ligado al Y)
|
Presencia
|
|||||||
Thompson, J.S y Thompson, M.W: (1980). Genética Médica. México: Salvat.
Richer, L.(2001).Genetica y Leyes.Peru.EdicionesGenovo.
No hay comentarios:
Publicar un comentario